Tyrannosaurus rex
"King of the tyrant lizards"
Sobre esta espécie
Tyrannosaurus rex is arguably the most famous dinosaur in the world. It lived at the end of the Cretaceous period, 68 to 66 million years ago, in what is now North America. At up to 12 meters long and over 8 metric tons, it was the apex predator of its ecosystem. Its serrated teeth measured up to 20 centimeters. While its tiny arms are often mocked, they were muscular enough to lift up to 200 kg. Recent research suggests T. rex may have had proto-feathers on parts of its body, as seen in close relatives.
Geological formation & environment
Tyrannosaurus rex is found mainly in the Hell Creek Formation, exposed in Montana, South Dakota, North Dakota, and Wyoming, and in the Frenchman Formation of Saskatchewan, Canada. Both date to the Upper Maastrichtian (~68–66 Ma). The environment was a warm subtropical alluvial plain crossed by meandering rivers and covered by dense forests of angiosperms, araucarias, and cycads. There were no polar ice caps and sea levels were higher than today. The ecosystem was dominated by large herbivores such as Triceratops and Edmontosaurus, which served as prey for T. rex. Other inhabitants included Ankylosaurus, Pachycephalosaurus, Dakotaraptor, crocodilians, and turtles. The formation is exceptional because periodic rivers rapidly buried carcasses, creating favorable conditions for fossilization.
Image gallery


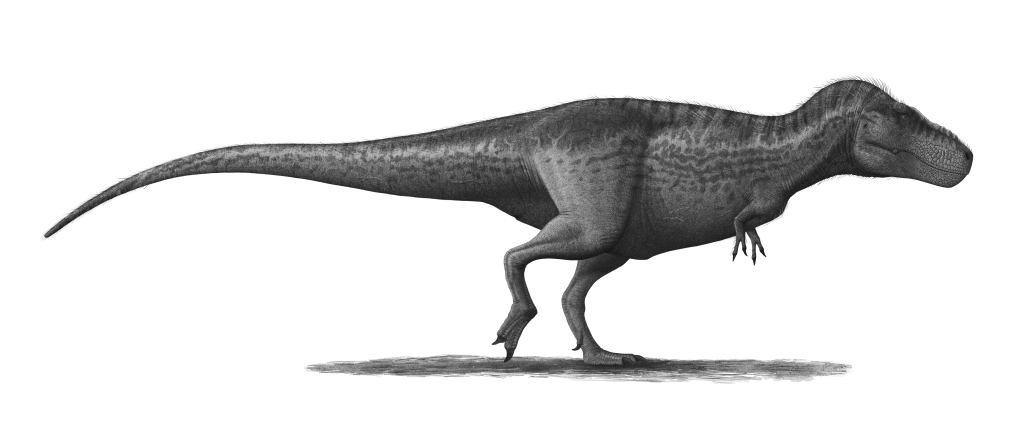





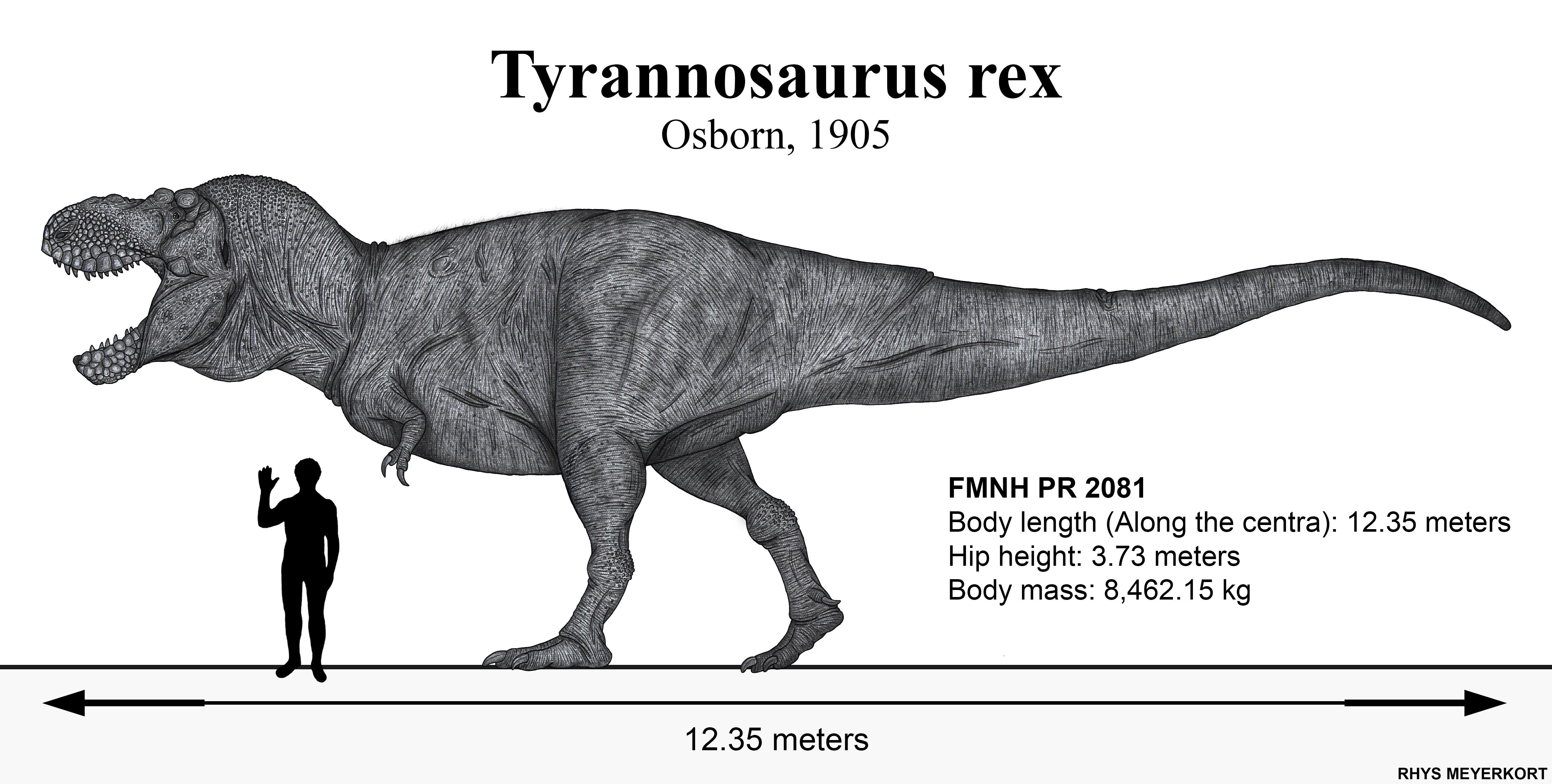




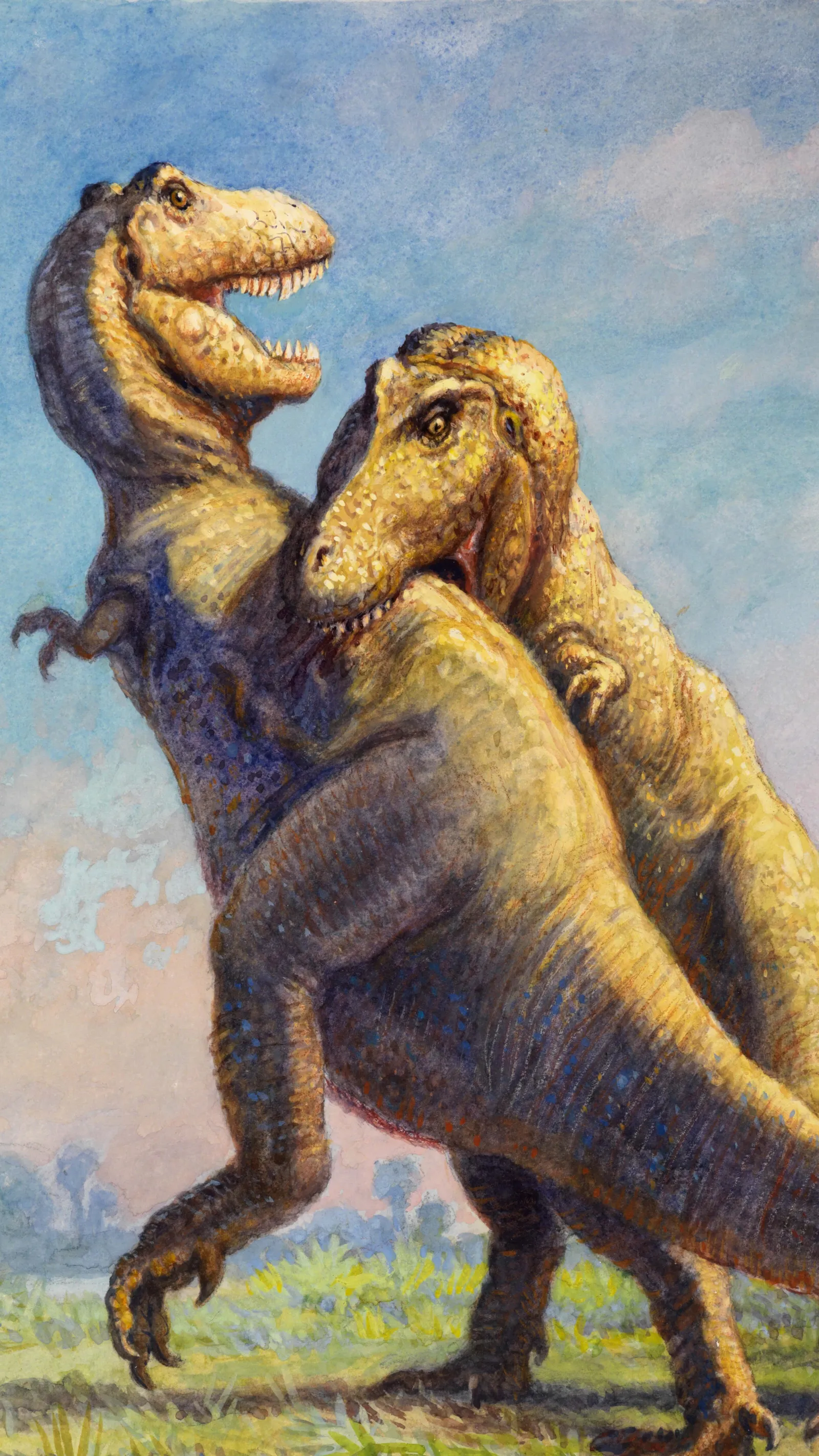


'Sue' (FMNH PR 2081) at the Field Museum, Chicago — the most complete (~90%) T. rex specimen ever found, discovered in 1990 by Sue Hendrickson.
Connie Ma, CC BY-SA 2.0
Ecology and behavior
Habitat
T. rex inhabited the coastal lowlands and humid tropical forests of the Hell Creek Formation, covering present-day Montana, South Dakota, Wyoming, and North Dakota 68–66 Ma ago. Climate was notably warmer: estimated mean annual temperature 11.3–11.6°C, with 191–197 cm/year of precipitation — similar to the modern Gulf Coast. The Western Interior Seaway created coastal plains, meandering rivers, and seasonally flooded swamps. Vegetation consisted of conifer forests (sequoias, cypresses, araucarias), fern meadows, tree ferns, and growing angiosperm diversity. Contemporary fauna included Triceratops horridus, Edmontosaurus regalis, Ankylosaurus magniventris, Pachycephalosaurus wyomingensis, Struthiomimus, pterosaurs, crocodilians, giant turtles, and primitive mammals like Didelphodon. Community analyses indicate T. rex represented roughly 24% of all large vertebrate biomass in the Hell Creek community (Scannella & Horner, 2010).
Feeding
T. rex was the absolute apex predator of the Hell Creek food chain. With an estimated bite force of 35,000–57,000 N — the greatest known for any land animal (Bates & Falkingham, 2012) — it could crush whole bones to access marrow, documented by deep bite marks on Triceratops and Edmontosaurus bones. The 2023 confirmation of scaly lips (Cullen et al., 2023) revealed that teeth were not permanently exposed as in crocodilians, but covered by a lip similar to monitor lizards. Primary prey were Triceratops and Edmontosaurus; healed bite marks on prey bones indicate some animals survived attacks. Hunting strategy was likely stealthy approach combined with explosive attack: adult T. rex could not sustain high-speed running (Hutchinson & Garcia, 2002), but used extraordinary smell and binocular vision to locate and ambush prey. Intraspecific cannibalism was documented by Longrich et al. (2010), who described T. rex bones bearing T. rex tooth marks.
Behavior and senses
T. rex was likely solitary as an adult. Adults display healed bite marks on skulls — direct evidence of violent intraspecific combat for territory or mates. The sensory system was among the most sophisticated of any dinosaur: Stevens (2006) calculated 55° binocular overlap — greater than any modern bird of prey — with visual acuity 13 times superior to humans. Olfactory bulbs were proportionally the largest of any known dinosaur, suggesting the ability to detect carcasses from kilometers away (Witmer & Ridgely, 2009). Carr et al. (2017) identified a crocodile-like facial sensory system — highly sensitive to touch, pressure, and temperature — possibly used for nest tending with thermal precision. Parental behavior is inferred by phylogenetic bracketing from crocodilians and birds, both of which guard nests and provide offspring care.
Physiology and growth
T. rex showed explosive growth during its juvenile phase: bone growth rings reveal adolescents gained more than 700 kg/year between ages 14–18 (Erickson et al., 2004) — one of the highest growth rates of any land vertebrate. It reached full adult size around age 20; maximum known longevity is ~28–30 years (Sue specimen: 28 growth rings). Wiemann et al. (2022) confirmed endothermic metabolism by analyzing nitrogen isotopes in bone collagen — intermediate between ectothermic reptiles and modern birds/mammals. Natural walking speed was ~4.6 km/h (fossilized trackways); estimated maximum speed was 17–25 km/h — true running would be biomechanically impossible for adults of 8+ metric tons (Hutchinson & Garcia, 2002). The forelimbs, despite being short (~1.2 m), had musculature capable of exerting ~200 kg of force with ~180° range, possibly used to stabilize carcasses during feeding.
Paleogeography
Continental configuration

Ron Blakey · CC BY 3.0 · Cretaceous, ~90 Ma
Fossil sites

Paleobiology Database · CC BY-SA 3.0
During the Maastrichtiano (~68–66 Ma), Tyrannosaurus rex inhabited Laramidia, the western half of present-day North America, separated from the east by the Western Interior Seaway, a shallow sea dividing the continent. The continents were in very different positions: India was drifting toward Asia, Antarctica was still connected to Australia, and South America was an isolated island.
Inventário de Ossos
Based on multiple specimens. The 'Sue' skeleton (Field Museum, Chicago), described in 2000, is ~90% complete and is the most complete T. rex ever found. The original holotype (AMNH 973) is at the American Museum of Natural History.

Found elements
Inferred elements
Scientific Literature
15 papers in chronological order — from the original description to recent research.
Tyrannosaurus and other Cretaceous carnivorous dinosaurs
Osborn, H.F. · Bulletin of the American Museum of Natural History
The founding paper of modern T. rex paleontology. Henry Fairfield Osborn describes specimen AMNH 973, collected by Barnum Brown from the Lance and Hell Creek formations of Wyoming, and establishes the genus and species Tyrannosaurus rex. Diagnostic characters defined include: colossal size (estimated ~12 m length), bipedal stance, extremely reduced forelimbs with two functional digits, laterally compressed serrated teeth up to 20 cm, and a large fenestrated skull of great robustness. Osborn compares the specimen with Allosaurus and with Cope's Manospondylus gigas (now recognized as T. rex), concluding it represents a new genus. The paper includes a description of hypothetical musculature and concludes T. rex was 'the most powerfully constructed carnivore yet discovered.' The fully upright posture depicted by Osborn — immortalized in Charles R. Knight's paintings for the AMNH — would be revised decades later, but the basic anatomical description remains a reference. This paper is the mandatory starting point for any research on the species.


Tyrannosaurus was not a fast runner
Hutchinson, J.R. & Garcia, M. · Nature
Seminal biomechanical study resolving one of the greatest controversies of 20th-century paleontology: could T. rex run? Hutchinson & Garcia build a computational musculoskeletal model based on comparative analysis with modern birds and crocodilians to estimate the minimum muscle mass needed for locomotion at different speeds. The result is definitive: for a 6,000 kg T. rex to run in the technical sense (with a flight phase, both feet off the ground), leg extensor muscles would need to represent ~86% of body mass — a biologically impossible value. Maximum sustainable speed is estimated at 17–25 km/h in fast walk. The authors note that juvenile T. rex, being proportionally lighter with different centers of mass, were likely significantly faster. The study ends decades of debate started by Robert Bakker in the 1970s, who envisioned T. rex as a fast runner. Fundamental ecological implication: the greatest land predator did not chase prey at high speed — it relied on ambush, stealth, and its superior sensory system.


Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull
Brochu, C.A. · Journal of Vertebrate Paleontology
The most comprehensive osteological monograph ever published on Tyrannosaurus rex, based primarily on the Sue specimen (FMNH PR 2081) — the most complete known, with ~90% of the skeleton preserved. Brochu provides systematic description of every skeletal element from skull to tail tip, and uses high-resolution computed tomography to reveal internal skull anatomy: elaborate pneumatic sinuses, neural canals, and cavities never previously documented. The work catalogs over 200 diagnostic characters and presents phylogenetic analysis placing tyrannosaurids as the sister group of Dromaeosauridae + Troodontidae within Coelurosauria. CT analysis reveals that skull bones were internally hollow — a structure combining lightness with resistance to extreme bite forces. Published as a 138-page Journal of Vertebrate Paleontology monograph, it is considered the definitive anatomical work on the species and the primary reference for all subsequent morphological studies.


Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs
Erickson, G.M. et al. · Nature
Erickson et al. cross-section bones of T. rex and other tyrannosaurids (Albertosaurus, Daspletosaurus, Gorgosaurus) and count annual growth rings — 'lines of arrested growth' (LAGs) — to determine specimen ages and reconstruct growth curves. The result is one of the most surprising findings of 21st-century paleontology: adolescent T. rex grew over 700 kg per year from ages 14–18 — a rate comparable only to large whales and fast-growing birds, completely outside the pattern of any living reptile. This explosive growth evidences endothermic ('warm-blooded') metabolism. T. rex reached full adult size (~5,800 kg) at ~20 years. Maximum documented longevity is 28 years, based on the Sue specimen. Comparative analysis with smaller tyrannosaurids reveals all followed the same accelerated growth pattern — suggesting elevated metabolism was a basal character of Tyrannosauridae.


Binocular vision in theropod dinosaurs
Stevens, K.A. · Journal of Vertebrate Paleontology
Stevens develops a method for reconstructing the visual fields of theropod dinosaurs in three dimensions, using digital skull endocasts and the geometry of eye orbits. Applied to T. rex, the result is unexpected with broad implications: 55° binocular overlap — substantially greater than any modern bird of prey (golden eagle: ~40°, peregrine falcon: ~35°) — and estimated visual acuity 13 times that of humans. This broad binocular field implies exceptional depth perception at long range, essential for precise ambushes. The study directly addresses Jack Horner's scavenger hypothesis, popularized in the media: animals relying primarily on carrion (vultures, hyenas) do not need refined binocular vision since they do not attack live prey. T. rex, with vision superior even to the best modern raptors, was almost certainly an active hunter. The paper also analyzes Troodon, Allosaurus, and other theropods, revealing binocular vision was a common evolutionary feature in predatory theropods.


Analyses of soft tissue from Tyrannosaurus rex suggest the presence of protein
Schweitzer, M.H. et al. · Science
In 2007, Mary Schweitzer and colleagues published a finding that shook paleontology: the identification of type I collagen fragments in cortical bone from a 68-million-year-old Tyrannosaurus rex. Using mass spectrometry, the team identified preserved peptide sequences without relying on genomic data. The discovery was initially met with skepticism — many researchers questioned whether the material could be microbial contamination or biofilm. Subsequent studies by Schweitzer's group and independent teams progressively confirmed the collagen's authenticity. Sequence analysis revealed that T. rex collagen is more similar to that of birds than crocodiles, providing independent molecular evidence for the phylogenetic position of dinosaurs as close relatives of birds. The work opened an entirely new field: palaeoproteomics. If proteins can survive 68 Ma under the right conditions, the molecular record of the past is far richer than imagined. The discovery also has implications for the search for biomolecules in other fossils and for understanding long-term preservation mechanisms. Published in Science, it has accumulated over 1,500 citations, becoming one of the most influential works in molecular palaeontology.
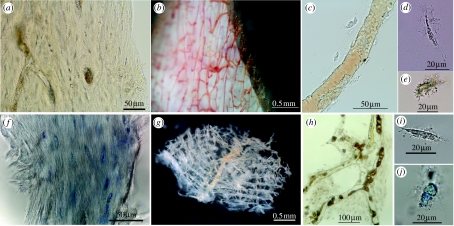
Cannibalism in Tyrannosaurus rex
Longrich, N.R., Horner, J.R., Erickson, G.M. & Currie, P.J. · PLOS ONE
Longrich et al. analyze T. rex skeletal material from the Hell Creek Formation (Montana) and identify T. rex tooth marks on T. rex bones — direct documentation of intraspecific cannibalism. Affected elements include metatarsals, tibias, and other long bones from multiple distinct individuals, all exhibiting scraping, perforation, and marrow extraction marks with morphology identical to that caused by T. rex on herbivore prey bones. The marks are inconsistent with combat injuries (wrong location, wrong angle, excessive depth) and most parsimonious as post-mortem alimentary cannibalism. There is no evidence that T. rex killed conspecifics to consume them, but there is clear evidence of feeding on their carcasses. The behavior may be: (1) opportunistic — when other food was scarce; (2) competitive — dominance over carcasses of individuals killed by other means; (3) pathological under environmental stress. T. rex joins a short list of non-avian dinosaurs with confirmed cannibalism.
Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics
Bates, K.T. & Falkingham, P.L. · Biology Letters
Bates & Falkingham apply two engineering methods to the T. rex skull to calculate maximum bite force: finite element analysis (FEA), which simulates mechanical stresses during biting, and multi-body dynamics, which models mandibular musculature. Results converge at 35,000–57,000 Newtons — the absolute record among all known land animals, fossil or living. For context: adult humans exert ~700 N; lions ~4,200 N; hyenas, specialized bone-crackers, ~4,500 N. T. rex could exert force 8–12 times greater than any living land predator. This capability explains the durophagous behavior documented in fossils: deep bite marks on solid Triceratops and Edmontosaurus bones indicate T. rex pulverized bones to extract fat-rich marrow. This food resource was inaccessible to other Hell Creek predators, giving T. rex a unique competitive advantage: the ability to extract calories from carcass parts all other animals were forced to ignore.


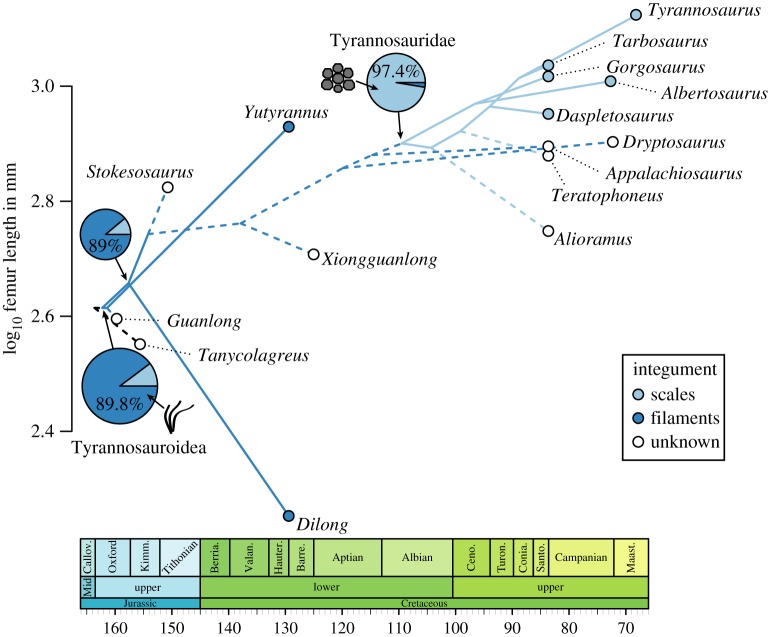
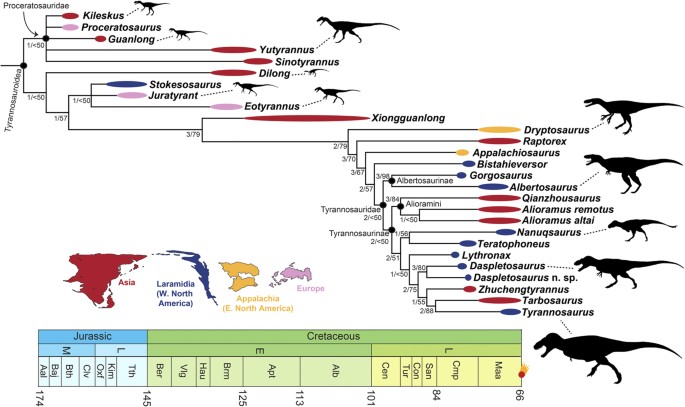
The phylogeny and evolutionary history of tyrannosauroid dinosaurs
Brusatte, S.L. & Carr, T.D. · Scientific Reports
In 2016, Steve Brusatte and Tom Carr published the most comprehensive phylogenetic analysis ever conducted for tyrannosauroids, including 26 taxa scored for 325 morphological characteristics. The work clarifies the evolutionary history of a group spanning more than 170 million years, from small Triassic ancestors to the iconic Late Cretaceous giants. Results identify five major subclades within Tyrannosauroidea and resolve the position of many controversial taxa, including Asian and European forms that were poorly understood. A central finding is that the gigantism of T. rex and its close relatives was a relatively late achievement in the group's history: tyrannosauroids were medium-sized animals for most of their existence, becoming dominant predators only after larger carnivores (such as allosaurids) went extinct in the Early Cretaceous. The analysis also documents how the group's diversification pattern tracked sea level fluctuations and continental fragmentation, especially the breakup of Laurasia. This work is now the standard reference for tyrannosauroid phylogeny.
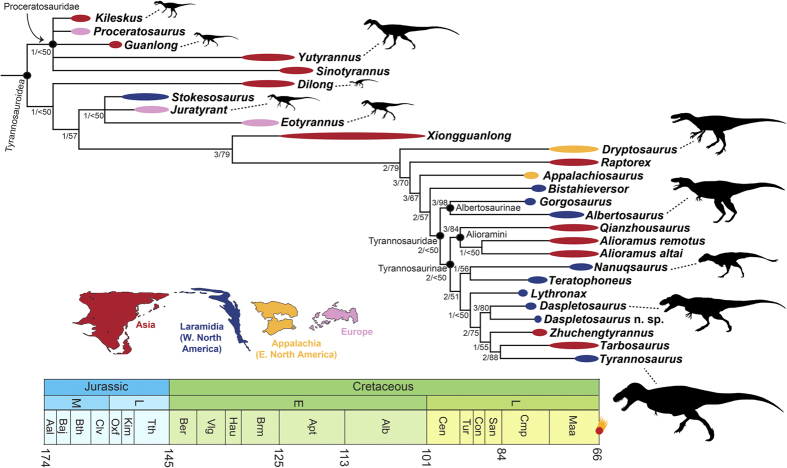
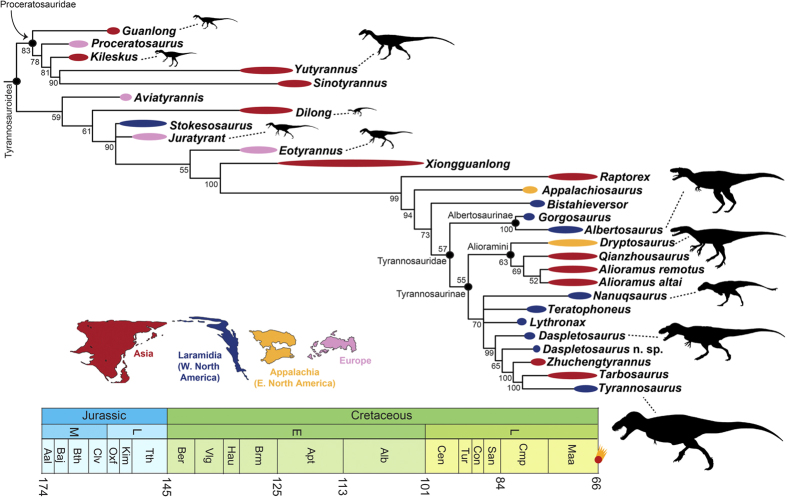
A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system
Carr, T.D. et al. · Scientific Reports
Carr and colleagues describe a new tyrannosaurid, Daspletosaurus horneri, from the Kirtlington Formation (Campanian), and identify in it — and by common ancestry in T. rex — a sophisticated facial sensory system with direct parallels in crocodilians. The snout surface presents numerous large neurovascular foramina and a rugose bone texture consistent with the presence of a densely innervated sensory integument, similar to the face of modern crocodilians. These receptors are highly sensitive to touch, pressure and temperature. The authors propose that this system allowed T. rex to: (1) build and repair nests precisely; (2) turn eggs without breaking them; (3) monitor incubation temperature; and (4) manipulate hatchlings gently. The implication is revolutionary for the cultural narrative of T. rex: the animal cinema portrayed as a relentless destroyer may also have been an attentive and caring parent. The work transforms our understanding of tyrannosaurid reproductive behavior and opens a new line of research on social intelligence in dinosaurs.
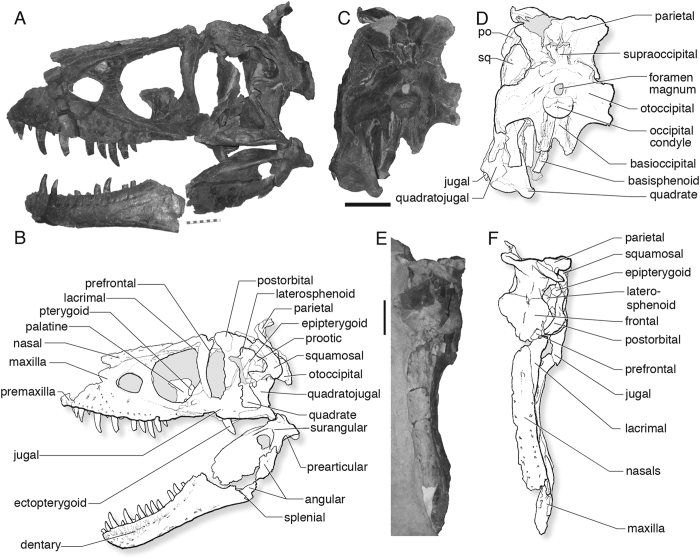
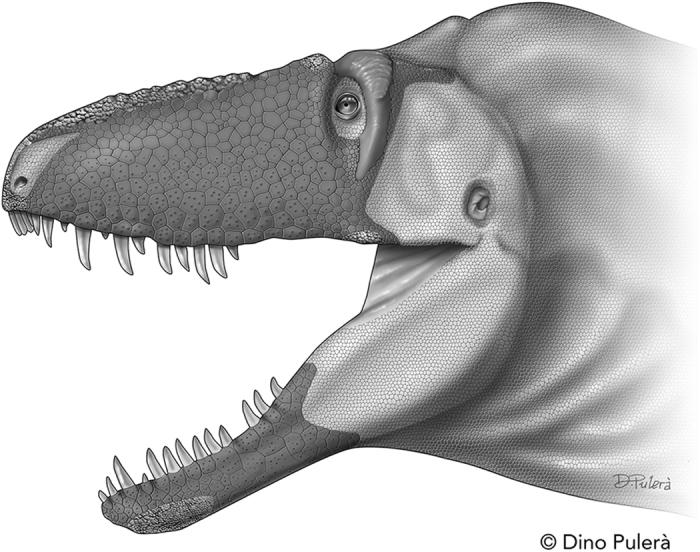
Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution
Bell, P.R. et al. · Biology Letters
One of the most debated questions about T. rex is whether it had feathers. In 2017, Phil Bell and colleagues published the first study based on actual skin impressions from the body of T. rex and other large tyrannosauroids. Analysis revealed small, non-overlapping scales covering at least the lateral and ventral surfaces. This contrasts with smaller, more ancient relatives like Yutyrannus huali (China, ~125 Ma), which clearly had filamentous coverings. Bell et al. propose two hypotheses: first, thermoregulation — large animals generate metabolic heat proportional to volume but dissipate it through skin proportional to surface area, making feathers counterproductive; second, the lineage may never have acquired body feathers at all. The work does not completely rule out feathers on unpreserved regions like the back, but makes extensive coverage unlikely. The discovery was widely reported and influenced T. rex depictions in later productions such as Jurassic World: Dominion (2022).

An older and exceptionally large adult specimen of Tyrannosaurus rex
Persons, W.S. et al. · The Anatomical Record
In 2019, Scott Persons and colleagues formally described specimen RSM P2523.8, nicknamed 'Scotty', from the Frenchman Formation (upper Maastrichtian) of Saskatchewan, Canada. Bone histology analysis revealed Scotty was one of the oldest individuals of T. rex ever found, estimated at around 28 years at death. More striking: the body mass estimate of ~8,800 kg surpasses all other known specimens, including Sue (~8,400 kg), making Scotty the largest confirmed T. rex. The study demonstrates that T. rex continued growing after sexual maturity, and that older individuals could significantly exceed the mass of younger ones. This has implications for understanding intraspecific variation and raises questions about the role of older individuals in T. rex ecology and behavior. The work also suggests the animal's maximum lifespan was greater than previous estimates indicated, opening the possibility of a late adult phase with distinct behaviors and ecological strategies.


The fast and the frugal: Divergent locomotory strategies drive limb lengthening in theropod dinosaurs
Dececchi, T.A. et al. · PLOS ONE
For decades, T. rex speed was hotly debated: was it an active hunter capable of catching fast prey, or a slow giant forced into scavenging? In 2020, Dececchi and colleagues published a comprehensive comparative biomechanical analysis spanning dozens of theropod species, separating two independent locomotory strategies: minimizing cost of transport (efficient long-distance walking) and maximizing top speed. For T. rex, the model indicates a preferred walking speed of 4.6–6.4 km/h, consistent with the 'metabolic economy' hypothesis: the animal was optimized for covering large distances efficiently, not for fast pursuit. This aligns with other computational biomechanics studies suggesting maximum speeds of 17-25 km/h for adult T. rex, well below early estimates of up to 45 km/h. The work also clarifies why long, gracile limbs evolved in different lineages for distinct reasons: speed in some groups, efficiency in others. For T. rex, the massive hindlimbs were adaptations for weight support and sustained locomotion, not sprinting.


Fossil biomolecules reveal an avian metabolism in the ancestral dinosaur
Wiemann, J. et al. · Nature
Wiemann et al. develop an innovative geochemical proxy for metabolism in fossils: the nitrogen isotopic composition of preserved bone collagen, which reflects the metabolic temperature during the animal's life. Applied to a phylogenetically diverse sample of Mesozoic archosaurs — including T. rex, Allosaurus, Triceratops, crocodilians, and modern birds — the method clearly discriminates ectotherms from endotherms. The result for T. rex is unambiguous: isotopic values are indistinguishable from modern birds and clearly separated from ectothermic reptiles. T. rex was endothermic, warm-blooded, with basal body temperature and metabolic rate comparable to mammals and birds. The study ends a debate lasting more than 50 years — since Robert Bakker proposed endothermic dinosaurs in 1969 — with direct evidence from the fossils themselves, not indirect inference. The finding is consistent with: the explosive growth documented by Erickson et al. (2004), the active behavior evidenced by Stevens (2006), and the role of active predator at the top of the food chain.


A reappraisal of the facial integument of theropod dinosaurs, with implications for the evolution of oral tissue in amniotes
Cullen, T.M. et al. · Science
A Science paper that fundamentally alters T. rex's appearance in art and popular culture. Cullen et al. use three independent lines of evidence: (1) phylogeny — crocodilians and lizards, the closest living relatives of dinosaurs, both possess lips covering the teeth; (2) dental morphology — T. rex teeth show a wear pattern consistent with labial coverage, unlike lipless crocodilian teeth; (3) mandibular morphology — the depth of the labial groove in jaw bones is consistent with soft tissue presence. The conclusion is that T. rex had scaly lips covering its teeth when the mouth was closed, like Varanidae lizards (monitors) — not permanently exposed teeth as in crocodilians or as depicted in Jurassic Park. The discovery has implications for bite force estimates, dental thermoregulation, and the evolutionary origin of the bird beak. Practically: all museum mounts and artistic reconstructions of T. rex with exposed teeth are technically incorrect.


Espécimes famosos em museus
Sue (FMNH PR 2081)
Field Museum of Natural History, Chicago
The most complete T. rex specimen ever found. Purchased at auction for US$ 8.36 million in 1997.

AMNH 5027
American Museum of Natural History, Nova York
The specimen that originated Osborn's formal 1905 description. One of the first T. rex specimens mounted for public display.
In cinema and popular culture
No prehistoric animal dominates the collective imagination like Tyrannosaurus rex. Its cinema debut came in 1925's The Lost World, an adaptation of Arthur Conan Doyle's novel, where it battled other giant reptiles in a scene that launched the monster movie tradition. In 1933, the T-rex made a brief appearance in King Kong, reinforcing the archetype of the invincible predator. In the golden age of animation, Fantasia (1940) immortalized the species in a sequence set to Stravinsky, portraying it as inevitable and relentless. The defining moment came in 1993 with Steven Spielberg's Jurassic Park, where the T-rex starred in scenes that would redefine science fiction for generations. The rain scene with the vibrating cup of water became a cultural icon. Despite being relatively accurate for its time, many details — such as movement-based vision and the absence of feathers — do not match current scientific consensus. The franchise expanded through five subsequent films, culminating in Jurassic World: Dominion (2022), each using the T-rex as a reliable narrative anchor for power and danger. Outside the Jurassic franchise, it appears in The Good Dinosaur (2015, Pixar), in a creative inversion where sauropods evolved intelligence and T-rexes took on the role of cattle ranchers. Cinematic representation solidified the image of a slow, visually-dependent biped; more recent science points to a far more dynamic animal, possibly with rudimentary plumage and acute senses.

Classificação
Descoberta
Curiosidade
T. rex had the most powerful bite of any land animal that ever lived: estimated at 35,000 to 57,000 Newtons, enough to crush entire bones to access the marrow.