Tyrannosaurus rex
"Rei dos lagartos tiranos"
Sobre esta espécie
O Tyrannosaurus rex é provavelmente o dinossauro mais famoso do mundo. Viveu no final do Cretáceo, há 68 a 66 milhões de anos, na América do Norte. Com até 12 metros de comprimento e mais de 8 toneladas, era o maior predador terrestre de seu ecossistema. Seus dentes, serrilhados como uma faca, mediam até 20 centímetros. Embora seus braços pequenos sejam motivo de piadas, eram musculosos o suficiente para levantar até 200 kg. Pesquisas recentes sugerem que o T. rex pode ter tido proto-penas em partes do corpo, como visto em parentes próximos.
Formação geológica e ambiente
O Tyrannosaurus rex é encontrado principalmente na Formação Hell Creek, que aflora em Montana, Dakota do Sul, Dakota do Norte e Wyoming, e na Formação Frenchman, no Canadá (Saskatchewan). Ambas datam do Maastrichtiano Superior (~68–66 Ma). O ambiente era uma planície aluvial de clima subtropical quente, cortada por rios meandrantes e coberta por florestas densas de angiospermas, araucárias e cicadófitos. Não havia calotas polares, e o nível do mar era mais alto que o atual. O ecossistema era dominado por herbívoros grandes como Triceratops e Edmontosaurus, que serviam de presa para o T. rex. Outros habitantes incluíam Ankylosaurus, Pachycephalosaurus, Dakotaraptor, crocodilos e tartarugas. A formação é excepcional porque os rios periódicos soterrava rapidamente carcaças, criando condições favoráveis para a fossilização.
Galeria de imagens


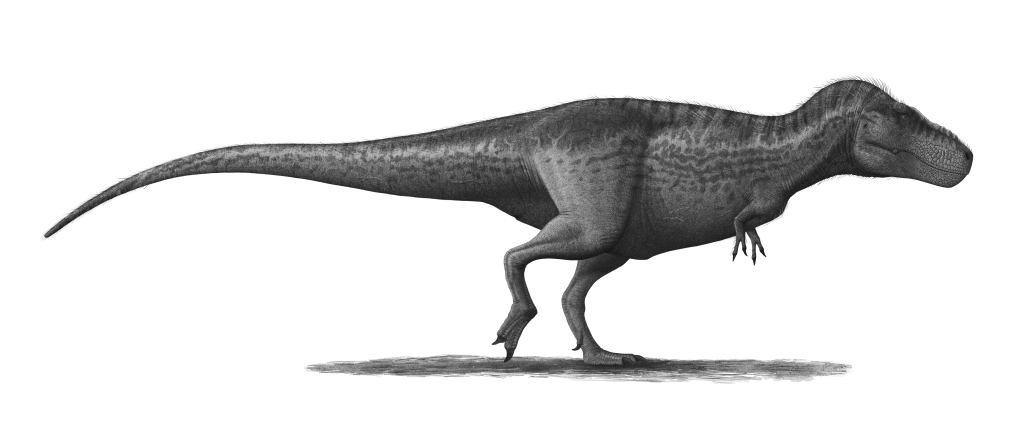





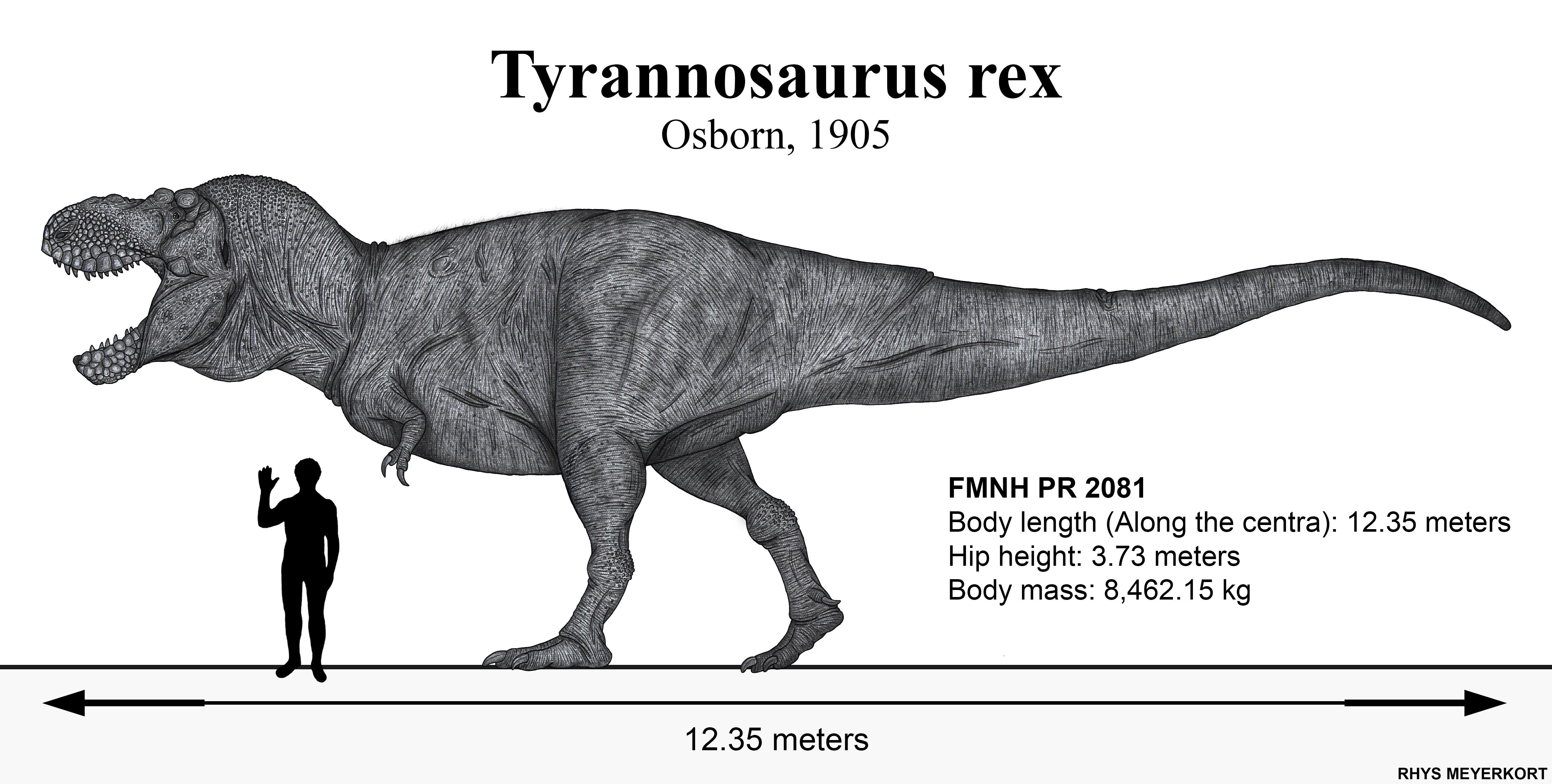




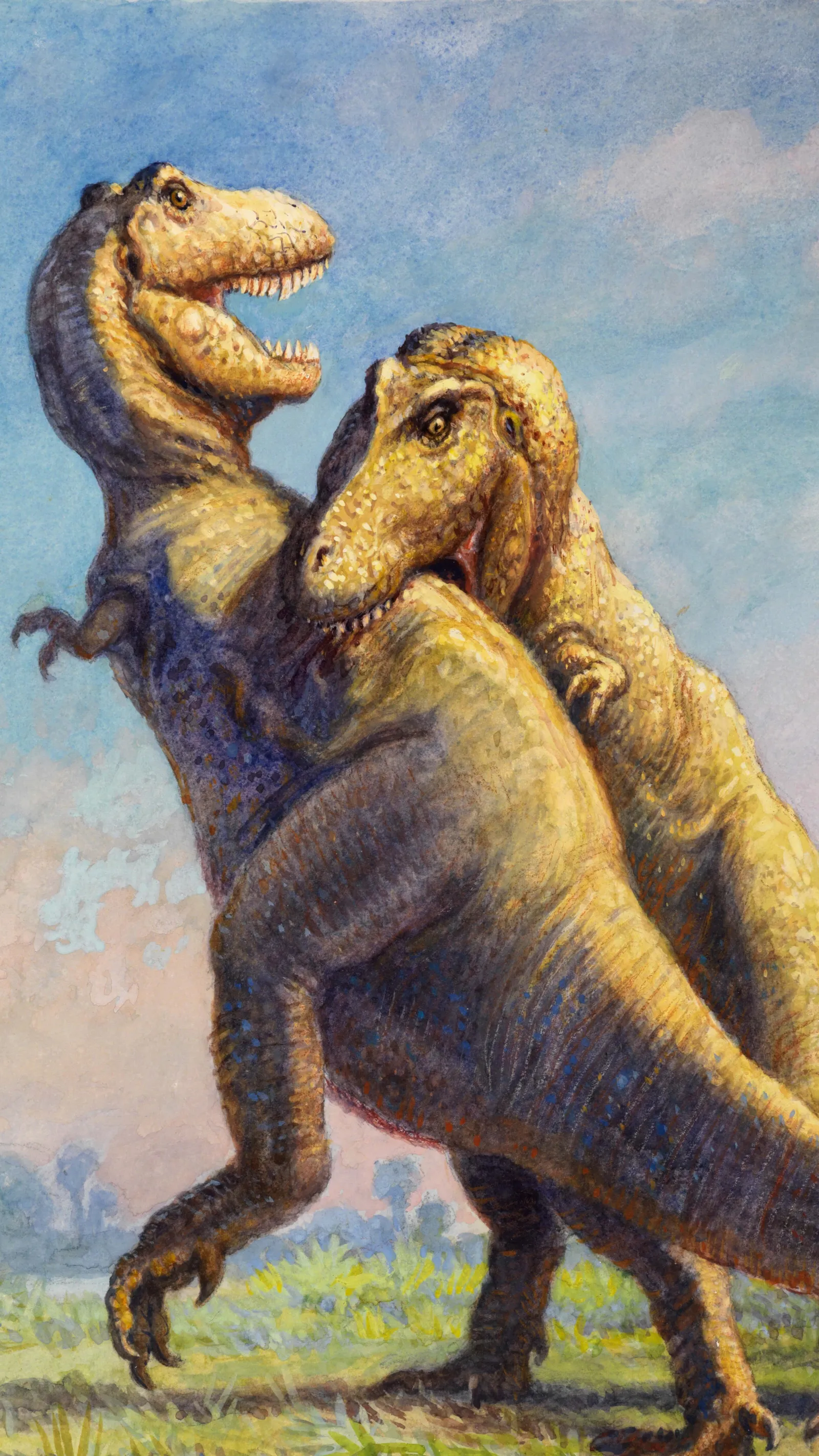


Esqueleto de 'Sue' (FMNH PR 2081) no Field Museum de Chicago — o espécime mais completo (~90%) de T. rex já encontrado, descoberto em 1990 por Sue Hendrickson.
Connie Ma, CC BY-SA 2.0
Ecologia e comportamento
Habitat
O T. rex habitou as planícies costeiras e florestas tropicais úmidas da Formação Hell Creek, que cobria o atual Montana, Dakota do Sul, Wyoming e Dakota do Norte há 68–66 Ma. O clima era notavelmente mais quente do que hoje: temperatura média anual estimada em 11,3–11,6°C, com precipitação de 191–197 cm/ano — semelhante ao atual litoral do Golfo do México. O Western Interior Seaway, um mar raso que dividia a América do Norte ao meio, criava planícies costeiras, rios meandrantes e brejos sazonalmente alagados. A vegetação era composta de florestas de coníferas (sequóias, ciprestes, araucárias), prados de samambaia, fetos arborescentes e crescente diversidade de angiospermas. A fauna contemporânea incluía Triceratops horridus, Edmontosaurus regalis, Ankylosaurus magniventris, Pachycephalosaurus wyomingensis, Struthiomimus, pterossauros, crocodilos, tartarugas gigantes e mamíferos primitivos como Didelphodon. Análises de comunidade indicam que o T. rex representava cerca de 24% de toda a biomassa de grandes vertebrados na Hell Creek (Scannella & Horner, 2010).
Alimentação
O T. rex era o predador no topo absoluto da cadeia alimentar de Hell Creek. Com força de mordida estimada em 35.000–57.000 N — a maior conhecida de qualquer animal terrestre (Bates & Falkingham, 2012) — era capaz de esmagar ossos inteiros para extrair o tutano, comportamento durofágico documentado por marcas de mordida profundas em ossos de Triceratops e Edmontosaurus. A confirmação de lábios escamosos em 2023 (Cullen et al., 2023) revelou que os dentes não ficavam permanentemente expostos como em crocodilos, mas cobertos por um lábio semelhante ao de lagartos monitor. As presas principais eram Triceratops e Edmontosaurus; marcas de mordida cicatrizadas em ossos indicam que algumas presas sobreviveram a ataques. A estratégia era provavelmente de aproximação furtiva combinada com ataque explosivo: o T. rex adulto não conseguia sustentar corrida prolongada (Hutchinson & Garcia, 2002), mas podia usar o olfato extraordinário e a visão binocular para localizar e emboscar presas. O canibalismo intra-específico foi documentado por Longrich et al. (2010), que descreveram ossos de T. rex com marcas de dentes de T. rex.
Comportamento e sentidos
O T. rex era provavelmente solitário na fase adulta. Os adultos apresentam marcas de mordida cicatrizadas nos crânios — evidência direta de combate intra-específico violento por território ou parceiros reprodutivos. O sistema sensorial era entre os mais sofisticados de qualquer dinossauro: Stevens (2006) calculou sobreposição binocular de 55° — superior à de qualquer ave de rapina moderna — com agudeza visual 13 vezes superior à humana. Os bulbos olfativos eram proporcionalmente os maiores de qualquer dinossauro conhecido, sugerindo capacidade de detectar carcaças a quilômetros de distância (Witmer & Ridgely, 2009). Carr et al. (2017) identificaram um sistema de sensores faciais semelhante ao dos crocodilos — altamente sensível a toque, pressão e temperatura — que pode ter sido utilizado para cuidar de ninhos com precisão térmica. O comportamento parental é inferido por analogia com crocodilos e aves, ambos os quais guardam ninhos e cuidam de filhotes.
Fisiologia e crescimento
O T. rex apresentava crescimento explosivo na fase juvenil: os anéis de crescimento ósseo revelam que adolescentes ganhavam mais de 700 kg/ano entre os 14 e 18 anos (Erickson et al., 2004) — uma das taxas de crescimento mais altas de qualquer vertebrado terrestre. Atingia o tamanho adulto pleno por volta dos 20 anos; o máximo de longevidade conhecida é de ~28–30 anos (espécime 'Sue': 28 anéis). Wiemann et al. (2022) confirmaram metabolismo endotérmico analisando isótopos de nitrogênio no colágeno ósseo — intermediário entre répteis ectotérmicos e aves/mamíferos modernos. A velocidade de caminhada natural era de ~4,6 km/h (pegadas fossilizadas); a velocidade máxima estimada era de 17–25 km/h — correr seria biomecanicamente impossível para adultos de 8+ toneladas (Hutchinson & Garcia, 2002). Os membros anteriores, apesar de curtos (~1,2 m), tinham musculatura capaz de exerter ~200 kg de força com amplitude de ~180°, possivelmente utilizados para estabilizar carcaças durante a alimentação.
Paleogeografia
Configuração continental

Ron Blakey · CC BY 3.0 · Cretáceo, ~90 Ma
Sítios fossilíferos

Paleobiology Database · CC BY-SA 3.0
Durante o Maastrichtiano (~68–66 Ma), Tyrannosaurus rex habitava a Laramídia, a metade ocidental do que hoje é a América do Norte, separada pelo Mar Interior do Oeste (Western Interior Seaway), um mar raso que dividia o continente ao meio. Os continentes estavam em posições muito diferentes das atuais: a Índia viajava em direção à Ásia, a Antártida ainda estava conectada à Austrália, e a América do Sul era uma ilha separada.
Inventário de Ossos
Baseado em múltiplos espécimes. O esqueleto 'Sue' (Field Museum, Chicago), descrito em 2000, é ~90% completo e é o mais completo já encontrado. O holótipo original (AMNH 973) está no American Museum of Natural History.

Estruturas encontradas
Estruturas inferidas
Literatura Científica
15 artigos em ordem cronológica — do artigo de descrição original até pesquisas recentes.
Tyrannosaurus and other Cretaceous carnivorous dinosaurs
Osborn, H.F. · Bulletin of the American Museum of Natural History
Artigo fundador da paleontologia moderna do T. rex. Henry Fairfield Osborn descreve o espécime AMNH 973, coletado por Barnum Brown nas formações Lance e Hell Creek do Wyoming, e estabelece o gênero e espécie Tyrannosaurus rex. Os caracteres diagnósticos definidos incluem: tamanho colossal (estimado em ~12 m de comprimento), postura bípede, membros anteriores extremamente reduzidos com dois dedos funcionais, dentes serrilhados lateralmente comprimidos de até 20 cm, e crânio fenestrado de grande robustez. Osborn compara o espécime com Allosaurus e com o Manospondylus gigas de Cope (cujo material é hoje reconhecido como T. rex), concluindo que se trata de um novo gênero. O trabalho inclui descrição da musculatura hipotética e conclui que o T. rex era 'o carnívoro mais poderoso jamais descoberto'. A postura completamente ereta representada por Osborn — e imortalizda nas pinturas de Charles R. Knight para o AMNH — seria revista décadas depois, mas a descrição anatômica básica permanece como referência. Este artigo é o ponto de partida obrigatório para qualquer pesquisa sobre a espécie.


Tyrannosaurus was not a fast runner
Hutchinson, J.R. & Garcia, M. · Nature
Estudo biomecânico seminal que resolve uma das maiores controvérsias da paleontologia do século XX: o T. rex conseguia correr? Hutchinson & Garcia constroem um modelo musculoesquelético computacional baseado em análise comparativa com aves e crocodilos modernos para estimar a massa muscular mínima necessária para locomoção em diferentes velocidades. O resultado é definitivo: para que um T. rex de 6.000 kg corresse no sentido técnico (com fase de voo, ambos os pés fora do chão), os músculos extensores da perna precisariam corresponder a ~86% da massa corporal — valor biologicamente impossível. A velocidade máxima sustentável é estimada em 17–25 km/h em marcha acelerada. Os autores ressaltam que juvenis de T. rex, por serem proporcionalmente mais leves e ter centros de massa diferentes, provavelmente eram significativamente mais rápidos. O estudo encerra décadas de debate iniciado por Robert Bakker nos anos 1970, que imaginava o T. rex como corredor veloz. Implicação ecológica fundamental: o maior predador terrestre não perseguia presas em alta velocidade — dependia de emboscada, furtividade e seu sistema sensorial superior.


Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull
Brochu, C.A. · Journal of Vertebrate Paleontology
A monografia osteológica mais abrangente já publicada sobre o Tyrannosaurus rex, baseada principalmente no espécime 'Sue' (FMNH PR 2081) — o mais completo conhecido, com ~90% do esqueleto preservado. Brochu realiza descrição sistemática de cada elemento esquelético, do crânio à ponta da cauda, e utiliza tomografia computadorizada de alta resolução para revelar a anatomia interna do crânio: seios pneumáticos elaborados, canais neurais e cavidades que nunca haviam sido documentadas. O trabalho cataloga mais de 200 características diagnósticas e apresenta análise filogenética posicionando os tiranossaurídeos como grupo-irmão de Dromaeosauridae + Troodontidae dentro de Coelurosauria. A análise CT revela que os ossos do crânio eram internamente ocos — estrutura que combinava leveza com resistência às forças de mordida extremas. Publicado como monografia do Journal of Vertebrate Paleontology com 138 páginas, é considerado o trabalho anatômico definitivo sobre a espécie e a referência primária para qualquer estudo morfológico posterior.


Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs
Erickson, G.M. et al. · Nature
Erickson et al. cortam transversalmente ossos de T. rex e outros tiranossaurídeos (Albertosaurus, Daspletosaurus, Gorgosaurus) e contam os anéis de crescimento anuais — as 'linhas de crescimento interrompido' (LAGs) — para determinar a idade dos espécimes e reconstruir as curvas de crescimento. O resultado é uma das descobertas mais surpreendentes da paleontologia do século XXI: o T. rex adolescente crescia mais de 700 kg por ano dos 14 aos 18 anos — uma taxa comparável apenas a grandes baleias e aves de crescimento rápido, totalmente fora do padrão de qualquer réptil vivo. Esse crescimento explosivo evidencia metabolismo endotérmico (de sangue quente). O T. rex atingia tamanho adulto pleno (~5.800 kg) aos ~20 anos. A longevidade máxima documentada é de 28 anos, baseada no espécime 'Sue'. A análise comparativa com os tiranossaurídeos menores revela que todos seguiam o mesmo padrão de crescimento acelerado — sugerindo que o metabolismo elevado era uma característica basal de Tyrannosauridae.


Binocular vision in theropod dinosaurs
Stevens, K.A. · Journal of Vertebrate Paleontology
Stevens desenvolve um método para reconstruir os campos visuais de dinossauros terópodes em três dimensões, utilizando moldagens digitais do crânio (endocastes) e a geometria das órbitas oculares. Aplicado ao T. rex, o resultado é inesperado e de amplas implicações: sobreposição binocular de 55° — substancialmente maior que qualquer ave de rapina moderna (águia dourada: ~40°, falcão-peregrino: ~35°) — e acuidade visual estimada em 13 vezes a humana. Esse campo binocular amplo implica percepção de profundidade excepcional a grande distância, essencial para emboscadas precisas. O estudo aborda diretamente a hipótese do T. rex necrófago, proposta por Jack Horner e popularizada na mídia: animais que dependem principalmente de carniça (abutres, hienas) não precisam de visão binocular refinada, pois não atacam presas vivas. O T. rex, com sua visão superior até às melhores aves de rapina modernas, quase certamente era caçador ativo. O artigo também analisa Troodon, Allosaurus e outros terópodes, revelando que visão binocular era uma característica evolutiva comum em terópodes predadores.


Analyses of soft tissue from Tyrannosaurus rex suggest the presence of protein
Schweitzer, M.H. et al. · Science
Em 2007, Mary Schweitzer e colegas publicaram uma descoberta que abalou a paleontologia: a identificação de fragmentos de colágeno tipo I em osso cortical de Tyrannosaurus rex com 68 milhões de anos. Usando espectrometria de massa, a equipe identificou sequências peptídicas preservadas sem recorrer a dados genômicos. A descoberta foi recebida com ceticismo inicial: muitos pesquisadores questionaram se o material poderia ser contaminação microbiana ou biopelícula bacteriana. Estudos subsequentes da própria Schweitzer e de grupos independentes confirmaram progressivamente a autenticidade do colágeno. A análise das sequências revelou que o colágeno de T. rex é mais similar ao de aves do que ao de crocodilos, fornecendo evidência molecular independente para a posição filogenética dos dinossauros como parentes próximos das aves. O trabalho abriu um campo inteiramente novo: a paleoproteômica. Se proteínas podem sobreviver 68 Ma em condições certas, o registro molecular do passado é muito mais rico do que se imaginava. A descoberta também tem implicações para a busca de biomoléculas em outros fósseis e para o entendimento dos mecanismos de preservação a longo prazo. A pesquisa foi publicada na revista Science e gerou mais de 1.500 citações, tornando-se um dos trabalhos mais influentes da paleontologia molecular.
Cannibalism in Tyrannosaurus rex
Longrich, N.R., Horner, J.R., Erickson, G.M. & Currie, P.J. · PLOS ONE
Longrich et al. analisam material esquelético de T. rex da Formação Hell Creek (Montana) e identificam marcas de dentes de T. rex em ossos de T. rex — documentação direta de canibalismo intra-específico. Os elementos afetados incluem metatarsos, tíbias e outros ossos longos de múltiplos indivíduos distintos, todos exibindo marcas de raspagem, perfuração e extração de tutano com morfologia idêntica à causada pelo T. rex em ossos de presas herbívoras. As marcas são inconsistentes com ferimentos de combate (localização errada, ângulo errado, profundidade excessiva) e mais parcimoniosas como canibalismo alimentar pós-morte. Não há evidência de que o T. rex matava coespecíficos para consumi-los, mas há evidência clara de que se alimentava de seus cadáveres. O comportamento pode ser: (1) oportunístico — quando outros alimentos escasseavam; (2) competitivo — dominância sobre carcaças de indivíduos mortos por outros meios; (3) patológico em condições de estresse ambiental. O T. rex se junta a uma curta lista de dinossauros não-aviários com canibalismo confirmado.
Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics
Bates, K.T. & Falkingham, P.L. · Biology Letters
Bates & Falkingham aplicam dois métodos de engenharia ao crânio do T. rex para calcular a força de mordida máxima: análise de elementos finitos (FEA), que simula as tensões mecânicas durante a mordida, e dinâmica de múltiplos corpos, que modela a musculatura mandibular. Os resultados convergem em 35.000–57.000 Newtons — o recorde absoluto entre todos os animais terrestres conhecidos, fósseis ou viventes. Para contextualizar: humanos adultos exercem ~700 N; leões ~4.200 N; hienas, especializadas em quebrar ossos, ~4.500 N. O T. rex podia exercer uma força 8–12 vezes maior que qualquer predador terrestre vivo. Essa capacidade explica o comportamento durofágico documentado nos fósseis: marcas de mordida profundas em ossos maciços de Triceratops e Edmontosaurus indicam que o T. rex trituravaos ossos para extrair o tutano rico em gordura. Esse recurso alimentar era inacessível para outros predadores de Hell Creek, dando ao T. rex uma vantagem competitiva única: a capacidade de extrair calorias de partes da carcaça que todos os outros animais eram obrigados a ignorar.


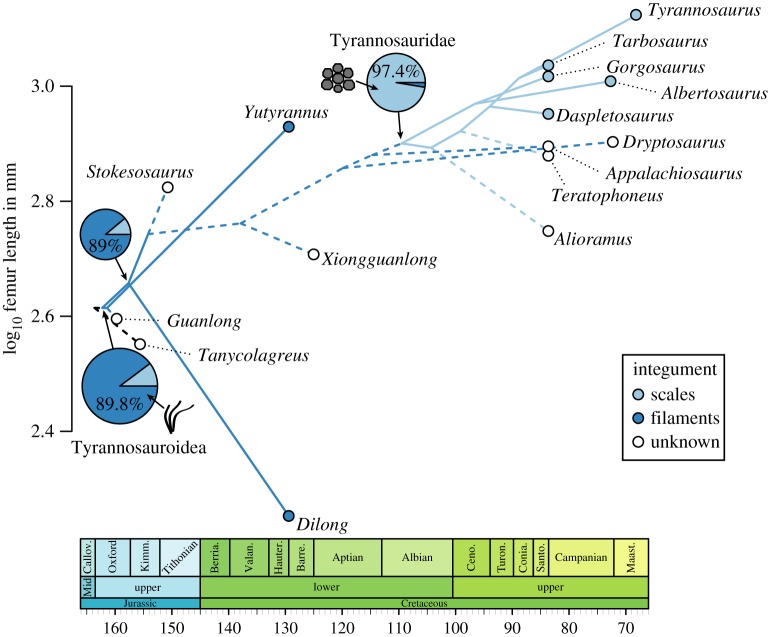
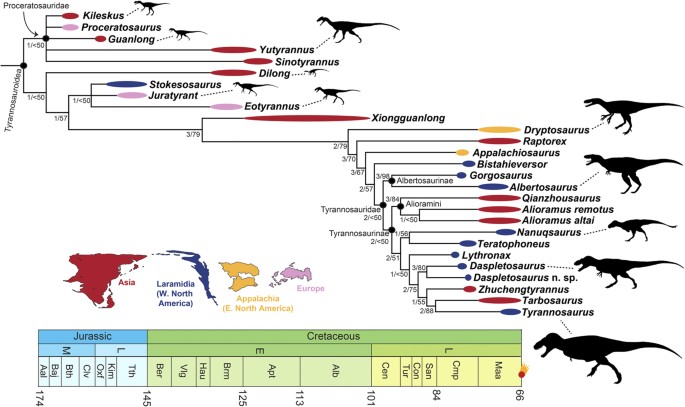
The phylogeny and evolutionary history of tyrannosauroid dinosaurs
Brusatte, S.L. & Carr, T.D. · Scientific Reports
Em 2016, Steve Brusatte e Tom Carr publicaram a análise filogenética mais abrangente já realizada para os tiranossaurídeos, incluindo 26 táxons avaliados para 325 características morfológicas. O trabalho esclarece a história evolutiva de um grupo com mais de 170 milhões de anos, desde pequenos ancestrais do Triássico até os gigantes icônicos do Cretáceo Superior. Os resultados identificam cinco grandes subclados dentro de Tyrannosauroidea e resolvem a posição de muitos táxons controversos, incluindo formas asiáticas e europeias que eram mal compreendidas. Um achado central é que o gigantismo de T. rex e seus parentes próximos foi uma conquista relativamente tardia na história do grupo: os tiranossaurídeos foram animais de médio porte por a maior parte de sua existência, tornando-se os predadores dominantes apenas quando os carnossauros maiores (como alosaurídeos) foram extintos no início do Cretáceo. A análise também documenta como o padrão de diversificação do grupo seguiu as flutuações do nível do mar e a divisão de continentes, especialmente a fragmentação da Laurásia. O trabalho é hoje a referência padrão para a filogenia do grupo.
A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system
Carr, T.D. et al. · Scientific Reports
Carr e colaboradores descrevem um novo tiranossaurídeo, Daspletosaurus horneri, da Formação Kirtlington (Campaniano), e identificam nele — e pelo método de ancestralidade comum no T. rex — um sistema de sensores faciais sofisticado com paralelo direto nos crocodilos. A superfície do focinho apresenta numerosos foramina neurovasculares grandes e uma textura óssea rugosa consistente com a presença de um tegumento sensorial denervado, semelhante ao da face de crocodilos modernos. Esses receptores são altamente sensíveis a toque, pressão e temperatura. Os autores propõem que esse sistema permitia ao T. rex: (1) construir e reparar ninhos com precisão; (2) virar ovos sem quebrá-los; (3) monitorar temperatura de incubação; e (4) manipular filhotes recém-nascidos com delicadeza. A implicação é revolucionária para a narrativa cultural do T. rex: o animal que o cinema retratou como destruidor implacável pode ter sido também um pai atento e cuidadoso. O trabalho transforma nossa compreensão do comportamento reprodutivo de tiranossaurídeos e abre uma nova linha de pesquisa sobre inteligência social em dinossauros.
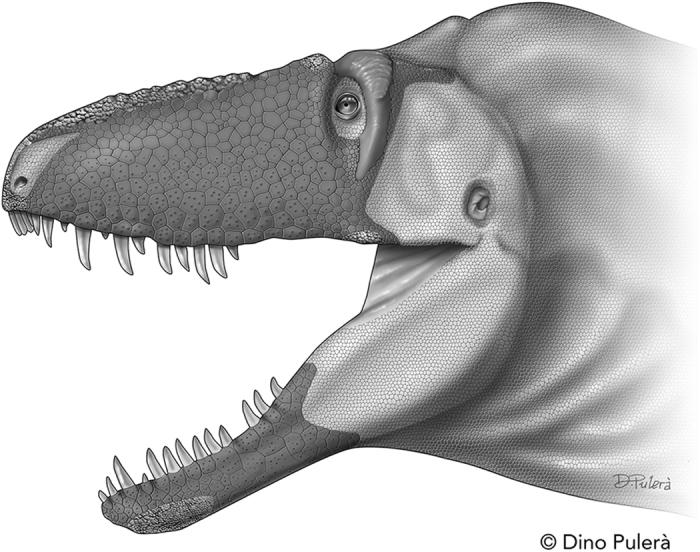
Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution
Bell, P.R. et al. · Biology Letters
Uma das questões mais debatidas sobre T. rex é se ele tinha penas. Em 2017, Phil Bell e colegas publicaram o primeiro estudo baseado em impressões de pele reais do corpo de T. rex e outros grandes tiranossaurídeos. A análise revelou escamas pequenas e não sobrepostas cobrindo ao menos as superfícies lateral e ventral do animal. Essa descoberta contrasta com parentes menores e mais antigos da linhagem, como Yutyrannus huali (China, ~125 Ma), que claramente possuíam coberturas filamentosas. Bell et al. propõem duas hipóteses para explicar a ausência de penas nos grandes tiranossaurídeos: a primeira é termorregulação, grandes animais geram calor metabólico proporcional ao volume mas dissipam pelo couro proporcional à superfície, então penas seriam contraproducentes; a segunda é que a linhagem nunca adquiriu penas no corpo, mesmo que parentes tenham adquirido. O trabalho não exclui completamente a presença de penas em regiões não preservadas como o dorso, mas torna improvável uma cobertura extensiva. A descoberta foi amplamente divulgada e influenciou a representação do T. rex em produções posteriores como Jurassic World: Dominion (2022).

An older and exceptionally large adult specimen of Tyrannosaurus rex
Persons, W.S. et al. · The Anatomical Record
Em 2019, Scott Persons e colegas descreveram formalmente o espécime RSM P2523.8, apelidado de 'Scotty', proveniente da Formação Frenchman (Maastrichtiano superior) de Saskatchewan, Canadá. A análise de histologia óssea revelou que Scotty era um dos indivíduos mais velhos já encontrados de T. rex, com estimativa de cerca de 28 anos na morte. Mais impactante ainda: a estimativa de massa corporal de ~8.800 kg supera a de todos os outros espécimes conhecidos, incluindo Sue (~8.400 kg), tornando Scotty o maior T. rex confirmado. O estudo demonstra que T. rex continuava crescendo após a maturidade sexual, e que os indivíduos mais velhos podiam superar significativamente a massa dos mais jovens. Isso tem implicações para a compreensão da variação intraespecífica da espécie e levanta questões sobre o papel dos indivíduos mais velhos na ecologia e comportamento de T. rex. O trabalho também sugere que a expectativa de vida máxima do animal era maior do que estimativas anteriores indicavam, abrindo espaço para uma fase adulta tardia com comportamentos e estratégias ecológicas distintas.


The fast and the frugal: Divergent locomotory strategies drive limb lengthening in theropod dinosaurs
Dececchi, T.A. et al. · PLOS ONE
Por décadas, a velocidade de T. rex foi alvo de debate: era um caçador ativo capaz de alcançar presas velozes, ou um gigante lento forçado ao carnivorismo? Em 2020, Dececchi e colegas publicaram uma análise biomecânica comparativa abrangendo dezenas de espécies de terópodes, separando duas estratégias locomotoras independentes: minimização do custo de transporte (andar eficiente por longas distâncias) e maximização da velocidade máxima. Para T. rex, o modelo indica uma velocidade de caminhada preferida de 4,6 a 6,4 km/h, coerente com a hipótese de 'economia metabólica': o animal era otimizado para percorrer grandes distâncias de forma energeticamente eficiente, não para perseguição veloz. Isso está alinhado com outros estudos de biomecânica computacional que sugerem velocidades máximas na faixa de 17-25 km/h para T. rex adulto, bem abaixo das estimativas iniciais de até 45 km/h. O trabalho também clarifica por que membros longos e gráceis evoluíram em diferentes linhagens por razões distintas: velocidade em alguns grupos, eficiência em outros. Para T. rex, os membros posteriores massivos eram adaptações para sustentar peso e locomoção sustentada, não para sprints.


Fossil biomolecules reveal an avian metabolism in the ancestral dinosaur
Wiemann, J. et al. · Nature
Wiemann et al. desenvolvem um proxy geoquímico inovador para metabolismo em fósseis: a composição isotópica do nitrogênio no colágeno ósseo preservado, que reflete a temperatura metabólica durante a vida do animal. Aplicado a uma amostra filogeneticamente diversa de arcossauros mesozoicos — incluindo T. rex, Allosaurus, Triceratops, crocodilos e aves modernas — o método discrimina claramente ectotérmicos de endotérmicos. O resultado para o T. rex é inequívoco: os valores isotópicos são indistinguíveis dos de aves modernas e claramente separados dos répteis ectotérmicos. O T. rex era endotérmico, de sangue quente, com temperatura corporal e taxa metabólica basais comparáveis a mamíferos e aves. O estudo encerra um debate que durou mais de 50 anos — desde que Robert Bakker propôs os dinossauros endotérmicos em 1969 — com uma evidência direta dos próprios fósseis, não inferência indireta. A descoberta é consistente com: o crescimento explosivo documentado por Erickson et al. (2004), a visão e comportamento ativo evidenciados por Stevens (2006), e o papel de predador ativo no topo da cadeia alimentar.


A reappraisal of the facial integument of theropod dinosaurs, with implications for the evolution of oral tissue in amniotes
Cullen, T.M. et al. · Science
Estudo publicado na Science que altera fundamentalmente a aparência do T. rex na arte e na cultura popular. Cullen et al. utilizam três linhas de evidência independentes: (1) filogenia — crocodilos e lagartos, os parentes vivos mais próximos dos dinossauros, ambos possuem lábios cobrindo os dentes; (2) morfologia dental — os dentes do T. rex apresentam padrão de desgaste consistente com cobertura labial, ao contrário dos dentes de crocodilos sem lábios; (3) morfologia da mandíbula — a profundidade do sulco labial nos ossos da mandíbula é consistente com a presença de tecido mole. A conclusão é que o T. rex tinha lábios escamosos que cobriam os dentes quando a boca estava fechada, como lagartos Varanidae (monitores) — não dentes permanentemente expostos como em crocodilos ou como retratado em Jurassic Park. A descoberta tem implicações para estimativas de força de mordida, termorregulação dental e para a evolução do bico das aves. Do ponto de vista prático: todas as montagens de museu e reconstruções artísticas do T. rex com dentes expostos estão tecnicamente incorretas.


Espécimes famosos em museus
Sue (FMNH PR 2081)
Field Museum of Natural History, Chicago
O espécime mais completo de T. rex já encontrado. Comprado em leilão por US$ 8,36 milhões em 1997.

AMNH 5027
American Museum of Natural History, Nova York
O espécime que originou a descrição formal de Osborn em 1905. Um dos primeiros T. rex montados para exibição pública.
No cinema e na cultura popular
Nenhum animal pré-histórico domina o imaginário coletivo como o Tyrannosaurus rex. Sua estreia no cinema ocorreu em 1925 em O Mundo Perdido, adaptação do romance de Arthur Conan Doyle, onde disputou espaço com outros répteis gigantes numa cena que inaugurou a tradição dos monstros do cinema. Em 1933, o T-rex apareceu brevemente em King Kong, reforçando o arquétipo do predador invencível. Na era clássica da animação, Fantasia (1940) imortalizou a espécie numa sequência ao som de Stravinsky, mostrando-o como um ser inevitável e implacável. O grande divisor de águas veio em 1993 com Jurassic Park, de Steven Spielberg, onde o T-rex protagonizou cenas que redefiniriam o gênero de ficção científica para gerações. A cena da chuva com o copo d'água vibrando tornou-se ícone cultural. Apesar da precisão relativa para a época, muitos detalhes como a visão baseada em movimento e a ausência de penas não correspondem ao consenso científico atual. A franquia se expandiu em cinco filmes subsequentes, até Jurassic World: Dominion (2022), cada um reutilizando o T-rex como âncora narrativa de segurança e poder. Fora da franquia Jurassic, aparece em O Bom Dinossauro (2015, Pixar), numa inversão criativa em que saurópodes evoluíram à inteligência e T-rexes assumiram o papel de pastores. A representação cinematográfica consolidou a imagem de um bípede lento e visual; a ciência mais recente aponta para um animal muito mais dinâmico, possivelmente com plumagem rudimentar e sentidos aguçados.

Classificação
Descoberta
Curiosidade
O T. rex tinha a mordida mais poderosa de qualquer animal terrestre que já existiu: estimada em 35.000 a 57.000 Newtons, suficiente para esmagar ossos inteiros para acessar o tutano.